How
we got here, and where we are going. Ever since the writings of Darwin and Huxley,
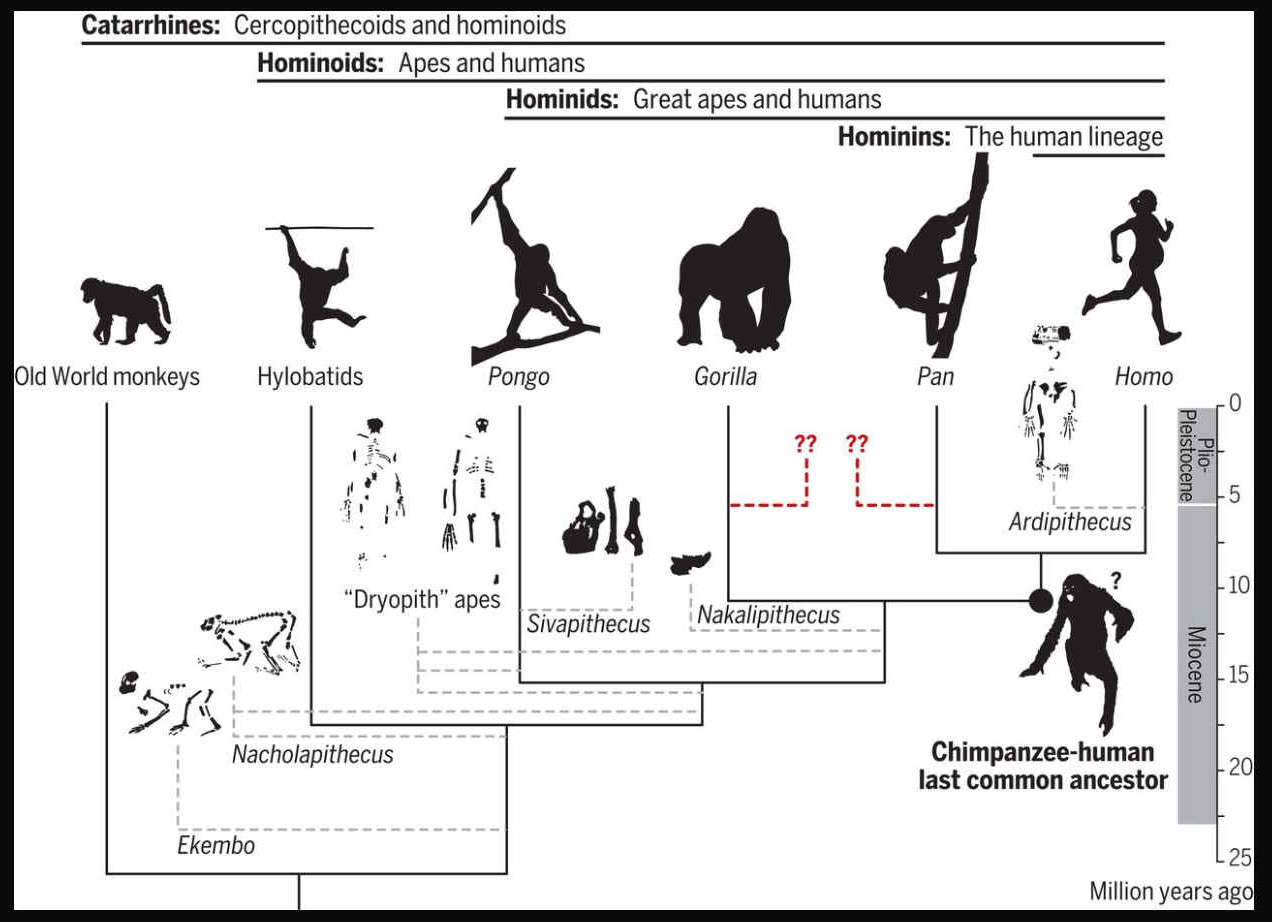
humans’ place in nature relative to apes (nonhuman hominoids) and the geographic origins of the human lineage (hominins) have been heavily debated. Humans diverged from apes [specifically, the chimpanzee lineage (Pan)] at some point between ~9.3 million and ~6.5 million years ago (Ma), and habitual bipedalism evolved early in hominins (accompanied by enhanced manipulation and, later on, cognition). To understand the selective pressures surrounding hominin origins, it is necessary to reconstruct the morphology, behavior, and environment of the Pan-Homo last common ancestor (LCA). “Top-down” approaches have relied on living apes (especially chimpanzees) to reconstruct hominin origins. However, “bottom-up” perspectives from the fossil record suggest that modern hominoids represent a decimated and biased sample of a larger ancient radiation and present alternative possibilities for the morphology and geography of the Pan-Homo LCA. Reconciling these two views remains at the core of the human origins problem.
Homo, is a Latin word meaning ‘human’ or ‘man’. The word neanderthalensis is based on the location where the first major specimen was discovered in 1856 – the Neander Valley in Germany. The German word for valley is ‘Tal’ although in the 1800s it was spelt ‘Thal’. Homo neanderthalensis therefore means ‘Human from the Neander Valley’.
Neanderthal, (Homo neanderthalensis, Homo sapiens neanderthalensis), also spelled Neandertal, member of a group of archaic humans who emerged at least 200,000 years ago during the Pleistocene Epoch (about 2.6 million to 11,700 years ago) and were replaced or assimilated by early modern human populations (Homo sapiens) between 35,000 and perhaps 24,000 years ago. Neanderthals inhabited Eurasia from the Atlantic regions of Europe eastward to Central Asia, from as far north as present-day Belgium and as far south as the Mediterranean and southwest Asia. Similar archaic human populations lived at the same time in eastern Asia and in Africa. Because Neanderthals lived in a land of abundant limestone caves, which preserved bones well, and where there has been a long history of prehistoric research, they are better known than any other archaic human group. Consequently, they have become the archetypal “cavemen.” The name Neanderthal (or Neandertal) derives from the Neander Valley (German Neander Thal or Neander Tal) in Germany, where the fossils were first found.
Until the late 20th century, Neanderthals were regarded as genetically, morphologically, and behaviorally distinct from living humans. However, more recent discoveries about this well-preserved fossil Eurasian population have revealed an overlap between living and archaic humans. Neanderthals lived before and during the last ice age of the Pleistocene in some of the most unforgiving environments ever inhabited by humans. They developed a successful culture, with a complex stone tool technology, that was based on hunting, with some scavenging and local plant collection. Their survival during tens of thousands of years of the last glaciation is a remarkable testament to human adaptation.
RITUALISTIC EXAMINATION
The common ancestor of hominins and chimpanzees existed around 6 Ma, while H. sapiens and our relatives, the Neanderthals (H. neanderthalensis), share a common ancestor who lived in the early Middle Pleistocene,
800-400 ka. Though our chimpanzee relatives continue to exist (somewhat precariously), Neanderthals disappeared approximately 40 000 years ago. Just as evaluation of chimpanzee cognition and behaviour can shed light on human origins, so too can comparisons between the archaeological record of near-contemporary H. sapiens and Neanderthals. What, then, do we know about the potential for ritual behaviours in our cousins? Let us first provide a sketch of what we know about their social and cognitive proclivities.
Having populated Europe and the Middle East between about 300,000 and 40,000 years BP, before being displaced by H. sapiens, Neanderthals left an
extensive - if patchy - record of their lifeways. Neanderthal groups employed various mobility strategies, and used a formal stone technology which represents an increase in hierarchical complexity over that of their Acheulean forebears. This stone technology was part of a wider techno-complex that included bone, claws, wood, shell and adhesive components, with tools appearing to be more diverse and task-specific (in some cases) than those of the Acheulean.
Neanderthal hunting strategies involved coordinated effort. For example, at Mauran (dating to MIS 3) in the foothills of the French Pyrenees, there is evidence that Neanderthal groups corralled migratory bison into natural geographical traps where they were slaughtered en masse, butchered and parts taken for consumption. This site was used for several hundred years, suggesting the maintenance of specialized, region-specific techniques, through the transmission of adaptive cultural knowledge, and an understanding of collective intentionality.
The presence of interregional variation in Neanderthal biface traditions similarly indicates the transmission of cultural knowledge between generations, with the technological continuity of the Mousterian a feature of the Eurasian Middle Palaeolithic. This technological stability, relative to near-contemporary H. sapiens, is the subject of debate, with recent research suggesting that a predominance of high-fidelity imitation without much experimentation in Neanderthal social learning may explain the technological stability. Neanderthals, then, were expert hunter–gatherers living in a variety of environments, who transmitted cultural knowledge over tens of thousands of years. But did they have the capacity for ritual, as we understand it in our own species?
Rituals for disposing of the dead are a significant part of the modern human experience, and intentional burials provide some of the clearest archaeological evidence for the presence of ritual. Chimpanzees have been known to place leafy branches on top of bodies of the deceased, though this behaviour is also performed for dead hetero-specifics, and might be a method for detecting movement. In hominins, intentional burial of the dead may date back to 400 000
BP - as suggested by the Iberian site of Sima de los Huesos - although, currently, evidence is only strong for the last 150 000 years. Indeed, the earliest undisputed evidence for burial is attributed to Neanderthal contexts. These burials typically occur in inhabited cave or rockshelter sites, which have been suggested to reflect an attachment to the dead and a desire to keep them physically and metaphorically close and safe after they have died. For example, at La Ferrassie (Dordogne, France) fetuses and young children were interred, possibly with grave goods (lithics).
IMPORTANT FOSSIL DISCOVERIES
The first Neanderthal fossil was found in 1829, but it was not recognised as a possible human ancestor until more fossils were discovered during the second half of the 19th century. Since then, thousands of fossils representing the remains of many hundreds of Neanderthal individuals have been recovered from sites across Europe and the Middle East. These include babies, children and adults up to about 40 years of age. As a result, more is known about this human ancestor than about any other.
KEY SPECIMENS:
- Le Moustier – a 45,000-year-old skull discovered in Le Moustier, France. The distinctive features of Neanderthals are already apparent in this adolescent individual. This shows that these characteristics were genetic and not developed during an individual’s lifetime.
- Shanidar 1 – upper jaw with teeth. The front teeth of Neanderthals often show heavy wear, a characteristic that is even found in young Neanderthals. It is probable that they used their teeth as a kind of vice to help them hold animal skins or other objects as they worked.
- La Ferrassie 1 – a 50,000-year-old skull discovered in 1909 in La Ferrassie, France. This skull of an elderly male has the features associated with ‘classic’ European Neanderthals.
- Amud 1 – a 45,000-year-old skull discovered in1961 by Hisashi Suzuki in Amud, Israel. This individual was more than 180 centimetres tall and had the largest brain of any fossil human (1740 cubic centimetres). Neanderthals probably migrated to the Middle East during times of harsh European winters. These individuals had less robust features than their European counterparts.
- Maba – a partial skull classifed as Homo sp. (species uncertain) and discovered in Maba, China. This partial skull, dated to about 120,000 – 140,000 years old, shows remarkable similarities to European Neanderthals and its discovery in southern China suggests the possibility that Neanderthals travelled further east than once thought. More fossil evidence from Asia is needed to understand the significance of this specimen.
- La Chapelle-aux-Saints – a 50,000-year-old skull discovered in 1908 in La Chapelle-aux-Saints, France. This male individual had lost most of his teeth and his skeleton showed evidence of major injuries and disease including a healed broken hip, and arthritis of the lower neck, back, hip and shoulders. He survived for quite some time with these complaints, which indicates that these people cared for the sick and elderly.
- Neanderthal 1 – a 45,000-year-old skullcap discovered in 1856 in Feldhofer Grotto, Neander Valley, Germany. This is the ‘type specimen’ or official representative of this species.
- Kebara 2 – 60,000-year-old partial skeleton discovered in 1983 in Kebara cave, Israel. This relatively complete skeleton belonged to an adult male. It was deliberately buried but as no grave goods were found it is difficult to infer any ritualistic behaviour.
- Lagar Velho – a 24,000-year-old skeleton of a Homo sapiens boy discovered in 1998 in Abrigo do Lagar Velho, central western Portugal. This specimen has been described by its discoverers (and particularly Eric Trinkhaus) as a Neanderthal-Homo sapiens hybrid. This interpretation was based on knee and leg proportions but as the head, pelvis and forearms are decidedly human it is more likely that the robustness is a climatic adaptation (see Tattersal and Schwartz). Comparisons to other humans of this period are difficult due to lack of knowledge on variations within child populations.
INTERBREEDING WITH MODERN HUMANS
Groundbreaking analysis of the Neanderthal genome (nuclear DNA and genes) published in 2010 shows that modern humans and Neanderthals did interbreed, although on a very limited scale. Researchers compared the genomes of five modern humans with the Neanderthal, discovering that Europeans and Asians share about 1-4% of their DNA with Neanderthals and Africans none. This suggests that modern humans bred with Neanderthals after moderns left Africa but before they spread to Asia and Europe. The most likely location is the Levant, where both species co-existed for thousands of years at various times between 50-90,000 years ago. Interestingly, the data doesn't support wide-scale interbreeding between the species in Europe, where it would have been most likely given their close proximity. Researchers are now questioning why interbreeding occurred on such a low scale, given that it was biologically possible. The answer may lie in cultural differences.
DENISOVIAN CAVE
An interesting case making headlines in 2010 was the discovery of a finger bone and tooth from Denisova cave in Russia. The bones were found in 2008 and date to about 30,000-50,000 years old. Mitochondrial DNA (mtDNA) was extracted from the remains, and then sequenced. The result was that the mtDNA did not match either modern human or Neanderthal mtDNA.
Little else could be gleaned from these studies so scientists started work on extracting nuclear DNA. This produced far more information. The 'Denisovians', as they have been nicknamed, were more closely related to Neanderthals than modern humans. This suggests the Neanderthals and 'Denisovans' shared a common ancestor after modern humans and Neanderthals split. Perhaps this ancestor left Africa half a million years ago with the Neanderthals spreading west to the Near East and Europe while the Denisovans headed east. However, this does not necessarily mean they are a 'new' species as they may be already known from fossils that have no DNA record to compare, such as Homo heidelbergensis or H. antecessor.
CANNIBALISM, DIET AND HUNTING
Great strides have been made in understanding Neanderthal hunting technology. Neanderthals were consummate hunters of medium and large-sized mammals. There is evidence that they used stone-tipped spears to hunt. For instance, it has been observed that Levallois points often bear impact scars on their tips (Shea 1988). In addition, retouched Mousterian points from several sites in western Europe (La Cotte de St. Brelade, Bouheben, Oscurusciuto, and Abric del Pastor) have been identified as bearing impact scars too (Villa & Soriano 2010). These stone-tipped spears were probably used at close-range for thrusting, or thrown short distances as part of an ambush hunting strategy — neither the stone points nor the features of Neanderthal anatomy support the notion that Neanderthals used long-distance projectiles (Shea & Sisk 2010, Churchill & Schmitt 2002, Rhodes & Churchill 2008).
There are several lines of evidence indicating that Neanderthals were capable hunters whose diets were dominated by meat. First is the faunal evidence from various sites, which indicates that they hunted and butchered red deer (Cervus elaphus), horse (Equus sp.), bison (Bison priscus), wooly rhinoceros (Coelodonta antiquitatis), and many other species of large and medium-sized ungulates (Patou-Mathis 2000). They focused on prime-aged adult prey, as opposed to juveniles and older adults, which are typically the target of cursorial predators (Stiner 2006). This behavior is likely to have begun 250,000 years ago or earlier, as seen at Hayonim Cave, Israel (Stiner 2005). There is also evidence that Neanderthals occasionally specialized in a single prey species, such as at Mauran, France, which is estimated to contain the remains of ~4,000 bison (Farizy et al. 1994). Similarly, the fauna at La Borde, France (Jaubert et al. 1990) and Wallertheim, Germany (Gaudzinski 1995) is also dominated by bovids, and these have been interpreted as kill sites. The site of Salzgitter-Lebenstedt (Germany) seems to have been a specialized reindeer-hunting site, where reindeer were taken in the early autumn during their annual migrations. The cut-marks clearly show meat processing, and the marrow-rich bones were also processed (Gaudzinski & Roebroeks 2000, Gaudzinski 2006). The evidence from all of these sites indicates that Neanderthals were able to hunt large game and often specialized in a species at a particular location and during a particular season.
Nitrogen and carbon isotope studies also support a Neanderthal diet focused on meat. Collagen obtained directly from at least a dozen Neanderthal bones from sites in France, Belgium, and Croatia reveal that Neanderthals were top-level carnivores who obtained most of their dietary protein from large herbivores (Richards & Trinkaus 2009, Bocherens 2009). Nevertheless, there is evidence that Neanderthals incorporated smaller animals and plant foods into their diets too. For instance, Neanderthals collected small game such as tortoises, shellfish, hares, rabbits, and birds in the Mediterranean region, and this trend increased dramatically just prior to or within the MP/UP transition (Stiner 2006). At the site of Vanguard Cave, Gibraltar, a small assemblage of mollusks was found, in addition to a few marine mammal bones (monk seal and dolphin) with cut-marks, indicating that Neanderthals exploited marine resources when they had the opportunity (Stringer et al. 2008).
There is also evidence that Neanderthals occasionally practiced cannibalism — at the site of Moula-Guercy, France, a number of Neanderthal remains were found with butchery marks indicating defleshing and marrow extraction. The similarity between the butchery marks on the Neanderthal bones and those on deer suggests the Neanderthals were consuming their dead for food (Defleur et al. 1999). Finally, there is evidence that Neanderthals consumed plant resources. Starch grains from legumes, date palms, and other plants — some of them exhibiting the transformations that happen during cooking — have been found preserved in dental calculus of Neanderthals from Shanidar, Iraq, and Spy, Belgium (Henry et al. 2011). In addition, a large assemblage of charred seeds and fruits (legumes, acorns, pistachios) were recovered from the Mousterian levels at Kebara Cave, Israel (Lev et al. 2005), and phytoliths recovered from sediments at Amud Cave suggest the consumption of grasses (Madella et al. 2002).
KANIS
REX
Man
took millions of years to develop from the apes, into
Homo Sapiens Sapiens, king of the primates until John
Storm was accidentally injected with a CRISPR
virus developed by Brazilian scientists working for the secret society; NeuWelt
Rittertum. The amateur anthropologist was later forced to
undo damaging alterations to his DNA, in so doing creating a new
species, physically and mentally superior to Homo S. Sapiens; named Homo
Sapiens Superior, or Kanis
Rex. These modifications were sufficient in scope to place him well
and truly in a new class. Progressing from Homo Sapiens Sapiens, to
become the king of men genetically.
The
modifications, only possible with the CyberCore Genetica™ super
computer, combined with the BioCore™
brain implant - and Hal's
AI,
were not just to his mental and muscular capabilities, but also to his
physical frame; his skeleton. Which is now stronger and lighter. All of
which in the natural world, would have taken a million or so years to
have achieved, assuming there would have been a clear natural selection advantage.

Sahelanthropus
Ardipithecus
Australopithecus
A. anamensis
A. afarensis
A. bahrelghazali
A. africanus
A. garhi
A. sediba
Kenyanthropus
Paranthropus robustus
Homo habilis
H. floresiensis
H. erectus
H. e. georgicus
H. cepranensis
H. antecessor
H. heidelbergensis
H. naledi
H. helmei
Homo neanderthalensis
Homo sapiens
H. s. idaltu
Homo Sapiens Sapiens
Homo
Sapiens Superior (Kanis
Rex)

EVOLUTION
Humans are apes (superfamily Hominoidea). The lineage of apes that eventually gave rise to humans first split from gibbons (family Hylobatidae) and orangutans (genus Pongo), then gorillas (genus Gorilla), and finally, chimpanzees and bonobos (genus Pan). The last split, between the human and
chimpanzee - bonobo lineages, took place around 8 - 4 million years ago, in the late Miocene epoch. During this split, chromosome 2 was formed from the joining of two other chromosomes, leaving humans with only 23 pairs of chromosomes, compared to 24 for the other apes. Following their split with chimpanzees and bonobos, the hominins diversified into many species and at least two distinct genera. All but one of these
lineages - representing the genus Homo and its sole extant species Homo
sapiens - are now extinct.
The genus Homo evolved from Australopithecus. Though fossils from the transition are scarce, the earliest members of Homo share several key traits with Australopithecus. The earliest record of Homo is the 2.8 million-year-old specimen LD 350-1 from Ethiopia, and the earliest named species are Homo habilis and Homo rudolfensis which evolved by 2.3 million years ago. H. erectus (the African variant is sometimes called H. ergaster) evolved 2 million years ago and was the first archaic human species to leave Africa and disperse across Eurasia. H. erectus also was the first to evolve a characteristically human body plan. Homo sapiens emerged in Africa around 300,000 years ago from a species commonly designated as either H. heidelbergensis or H. rhodesiensis, the descendants of H. erectus that remained in Africa. H. sapiens migrated out of the continent, gradually replacing or interbreeding with local populations of archaic humans.
Humans began exhibiting behavioral modernity about 160,000-70,000 years ago, and possibly earlier.
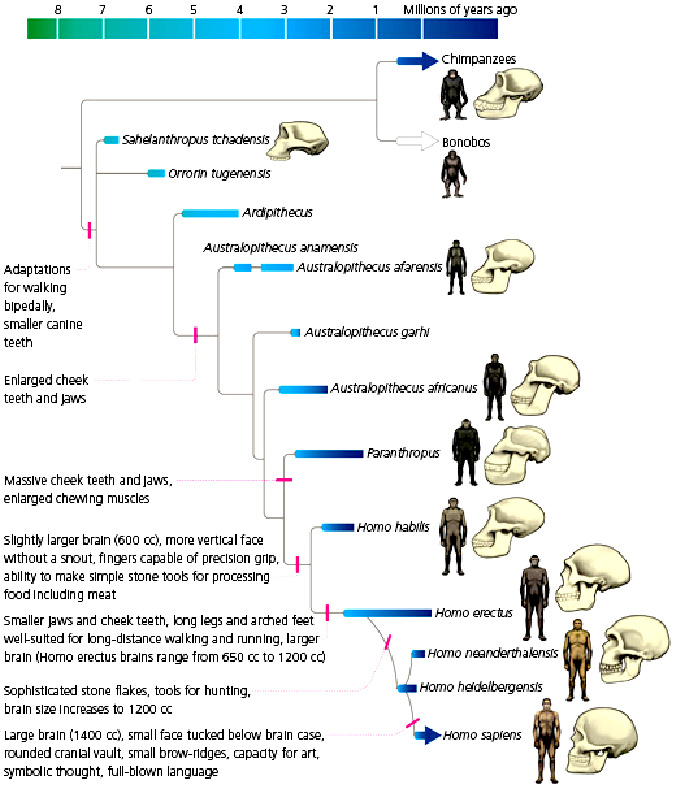
The "out of Africa" migration took place in at least two waves, the first around 130,000 to 100,000 years ago, the second (Southern Dispersal) around 70,000 to 50,000 years ago. H. sapiens proceeded to colonize all the continents and larger islands, arriving in Eurasia 125,000 years ago, Australia around 65,000 years ago, the Americas around 15,000 years ago, and remote islands such as Hawaii, Easter Island, Madagascar, and New Zealand between the years 300 and 1280 CE.
Human evolution was not a simple linear or branched progression but involved interbreeding between related species. Genomic research has shown that hybridization between substantially diverged lineages was common in human evolution. DNA evidence suggests that several genes of Neanderthal origin are present among all non sub-Saharan African populations, and Neanderthals and other hominins, such as Denisovans, may have contributed up to 6% of their genome to present-day non sub-Saharan African humans.
Human evolution is characterized by a number of morphological, developmental, physiological, and behavioral changes that have taken place since the split between the last common ancestor of humans and chimpanzees. The most significant of these adaptations are obligate bipedalism, increased brain size and decreased sexual dimorphism (neoteny). The relationship between all these changes is the subject of ongoing debate.

The
Cup of Christ is the Holy Grail, that has never been found, in all
searches through the ages.